01.Dunnill. MS, Massarella, GR, Anderson JA: A comparison of the quantitative anatomy of the bronchi in normal subjects, in status asthmaticus, in chronic bronchitis, and in emphysema. Thorax 1969; 24:176-179.
02.Jeffery PK. Pathology of asthma. In: TJH Clark, S Godfrey, TH Lee, NC Thompson. Asthma. London: Arnold; 2000:175-196.
03.Kradin, RL. Understanding Pulmonary Pathology. London. Elsevier: 2017.
04.Papi A, Brightling C, Pedersen SE, Reddel HK. Asthma. Lancet 2018; 391:783-800.
05.Roche WR, Beasley R, Williams JH, Holgate ST. Subepithelial fibrosis in the bronchi of asthmatics. Lancet 1(8637):520-4.
06.Latinen A, Altraja A, Kampe M, Linden M, Virtanen I, Laitinen LA. Tenascin is increased in airway basement membrane of asthmatics and decreased by an inhaled steroid. Am J Respir Crit Care Med 1997; 156:951-8.
07.Altraja A, Latinen A, Virtanen I et al. Expression of laminins in airways in various types of asthmatics patients: a morphometric study. Am J Respir Cell Mol Biol 1996; 15:482-8.
08.Sanerkin NG, Evans MD. The sputum in bronchial asthma: pathognomonic patterns. J Pathol Bacteriol 1965; 89:535 41.
09.Curschmann H: Uber Bronchiolitis exsudatira und ihr Verhaltuis zum Asthma nervosum. Dtsch Arch Klin Med 1883; 32:1-34.
10.Weller PF, Bach DS, Austen KF: Biochemical characterization of human eosinophil Charcot-Leyden crystal protein (lysophospholipase). J Biol Chem 1984; 259:15100-5.
11.Persson EK, Verstraete K, Heyndrickx I, Gevaert E, Aegerter H, Percier JM, Deswarte K, Verschueren KHG, Dansercoer A, Gras D, Chanez P, Bachert C, Gonçalves A, Van Gorp H, De Haard H, Blanchetot C, Saunders M, Hammad H, Savvides SN, Lambrecht BN. Protein crystallization promotes type 2 immunity and is reversible by antibody treatment. Science 2019; 364(6442):eaaw4295.
12.Charcot JM, Robin C. Observation de leucocythemie. Mem. Soc. Biol 1853; 5:44–50.
13.Leyden E. Zur Kenntniss des Bronchial-Asthma. Arch Pathol Anat Physiol Klin Med 1872; 54:32444.
14.Aikawa T, Shimura S, Sasaki H, Ebina M, Takishima T. Marked goblet cell hyperplasia with mucus accumulation in the airways of patients who died of severe acute asthma attack. Chest 1992; 101:916-21.
15.Ordoñez CL, Khashayar R, Wong HH, Ferrando R, Wu R, Hyde DM, Hotchkiss JA, Zhang Y, Novikov A, Dolganov G, Fahy JV. Mild and moderate asthma is associated with airway goblet cell hyperplasia and abnormalities in mucin gene expression. Am J Respir Crit Care Med 2001; 163:517-23.
16.Tavakoli S, Levine SJ, Shelhamer JH. Airway Mucus Secretion. In: Barnes PJ. et. al, eds. Asthma. Philadelphia: Lippincott-Raven, 1997:843-857.
17.Liu YC, Khawaja AM, Rogers DF. – Pathology of Airway Mucus Secretion in Asthma. In: Peter J. Barnes, Ian W. Rodger and Neil C. Thomson. Asthma. Basic Mechanisms and Clinical Management. San Diego: Academic Press; 1998: 205-227.
18.Liegeois MA, Hsieh A, Al-Fouadi M, Charbit AR, Yang CX, Hackett TL, Fahy JV. Cellular and molecular features of asthma mucus plugs provide clues about their formation and persistence. J Clin Invest 2025; 135:e186889.
19.Morgan LE, Jaramillo AM, Shenoy SK, Raclawska D, Emezienna NA, et al. 2021. Disulfide disruption reverses mucus dysfunction in allergic airway disease. Nat Commun 2021; 12:249.
20.Hogg JC. The pathology of asthma. J. Pathol Microbiol Immunol 1997; 105:735-−−45.
21.Bonser LR, Zlock L, Finkbeiner W, Erle DJ. Epithelial tethering of MUC5AC-rich mucus impairs mucociliary transport in asthma. J Clin Invest 2016; 126:2367-71.
22.Dunican EM, Watchorn DC, Fahy JV. Autopsy and Imaging Studies of Mucus in Asthma. Lessons Learned about Disease Mechanisms and the Role of Mucus in Airflow Obstruction. Ann Am Thorac Soc 2018; 15:S184-S191.
23.Aegerter H, Lambrecht BN. The Pathology of Asthma: What Is Obstructing Our View? Annu Rev Pathol 2023; 18:387-409.
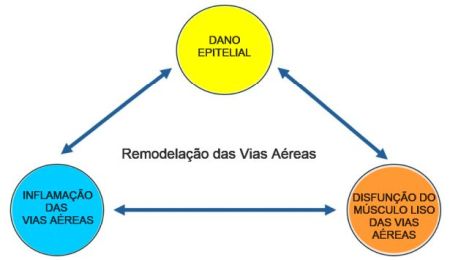
24.Tiotiu A, Steiropoulos P, Novakova S, Nedeva D, Novakova P, Chong-Neto H, Fogelbach GG, Kowal K. Airway Remodeling in Asthma: Mechanisms, Diagnosis, Treatment, and Future Directions. Arch Bronconeumol 2025; 61:31-40.
25.Bousquet J, Jeffery PK, Busse WW, Johnson M, Vignola AM. Asthma. From bronchoconstriction to airways inflammation and remodeling. Am J Respir Crit Care Med 2000; 161:1720-45.
26.Fehrenbach H, Wagner C, Wegmann M. Airway remodeling in asthma: what really matters. Cell Tissue Res 2017; 367:551-569.
27.Tang ML, Wilson JW, Stewart AG, Royce SG. Airway remodelling in asthma: current understanding and implications for future therapies. Pharmacol Ther 2006; 112:474-88.
28.Hough KP, Curtiss ML, Blain TJ, Liu RM, Trevor J, Deshane JS, Thannickal VJ. Airway Remodeling in Asthma. Front Med (Lausanne). 2020; 7:191.
29.James AL, Wenzel S. Clinical relevance of airway remodelling in airway diseases. Eur Respir J 2007; 30:134-55.
30.Thomas D, McDonald VM, Pavord ID, Gibson PG. Asthma remission: what is it and how can it be achieved? Eur Respir J 2022; 60:2102583.
31.Brightling CE, Gupta S, Gonem S, Siddiqui S. Lung damage and airway remodelling in severe asthma. Clin Exp Allergy 2012; 42:638-49.
32.Krings JG, Goss CW, Lew D, Samant M, McGregor MC, Boomer J, Bacharier LB, Sheshadri A, Hall C, Brownell J, Schechtman KB, Peterson S, McEleney S, Mauger DT, Fahy JV, Fain SB, Denlinger LC, Israel E, Washko G, Hoffman E, Wenzel SE, Castro M; National Heart, Lung, and Blood Institute's Severe Asthma Research Program Investigators. Quantitative CT metrics are associated with longitudinal lung function decline and future asthma exacerbations: Results from SARP-3. J Allergy Clin Immunol 2021; 148:752-762.
33. Calvén J, Ax E, Rådinger M. The Airway Epithelium-A Central Player in Asthma Pathogenesis. Int J Mol Sci 2020; 21:8907.
34.Heijink IH, Kuchibhotla VNS, Roffel MP, Maes T, Knight DA, Sayers I, Nawijn MC. Epithelial cell dysfunction, a major driver of asthma development. Allergy 2020; 75:1902-1917.
35.Olczyk P, Mencner L, Komosinska-Vassev K. The role of the extracellular matrix components in cutaneous wound healing. Biomed Res Int 2014;2014:747584.
36.Mostaço-Guidolin LB, Osei ET, Ullah J, Hajimohammadi S, Fouadi M, Li X, Li V, Shaheen F, Yang CX, Chu F, Cole DJ, Brandsma CA, Heijink IH, Maksym GN, Walker D, Hackett TL. Defective Fibrillar Collagen Organization by Fibroblasts Contributes to Airway Remodeling in Asthma. Am J Respir Crit Care Med 2019; 200:431-443.
37.Reeves SR, Kolstad T, Lien TY, Elliott M, Ziegler SF, Wight TN, Debley JS. Asthmatic airway epithelial cells differentially regulate fibroblast expression of extracellular matrix components. J Allergy Clin Immunol 2014; 134:663-670.e1.
38.Fang CL, Yin LJ, Sharma S, Kierstein S, Wu HF, Eid G, Haczku A, Corrigan CJ, Ying S. Resistin-like molecule-β (RELM-β) targets airways fibroblasts to effect remodelling in asthma: from mouse to man. Clin Exp Allergy 2015; 45:940-952.
39.Roche WR, Beasley R, Williams JH, Holgate ST. Subepithelial fibrosis in the bronchi of asthmatics. Lancet 1989; 1:520-4.
40.Mitchell PD, O'Byrne PM. Epithelial-derived cytokines in asthma. Chest 2017; 151:1338–44.
41.Porsbjerg CM, Sverrild A, Lloyd CM, et al. Anti-alarmins in asthma: targeting the airway epithelium with next-generation biologics. Eur Respir J 2020; 56:2000260 [https://doi.org/10.1183/ 13993003.00260-2020].
42.Bartemes KR, Kita H. Dynamic role of epithelium-derived cytokines in asthma. Clin Immunol 2012; 143:222-35.
43. Yang D, Han Z, Oppenheim JJ. Alarmins and immunity. Immunol Rev 2017; 280:41-56.
44. Cao L, Liu F, Liu Y, Liu T, Wu J, Zhao J, Wang J, Li S, Xu J, Dong L. TSLP promotes asthmatic airway remodeling via p38-STAT3 signaling pathway in human lung fibroblast. Exp Lung Res 2018; 44:288-301.
45. Wu J, Liu F, Zhao J, Wei Y, Lv J, Dong F, Bi W, Wang X, Wang J, Liu W, Dong L, Tian H. Thymic stromal lymphopoietin promotes asthmatic airway remodelling in human lung fibroblast cells through STAT3 signalling pathway. Cell Biochem Funct 2013; 31:496-503.
46.Jin A, Tang X, Zhai W, Li Y, Sun Q, Liu L, Yang X, Ren H, Lu S. TSLP-induced collagen type-I synthesis through STAT3 and PRMT1 is sensitive to calcitriol in human lung fibroblasts. Biochim Biophys Acta Mol Cell Res 2021; 1868:119083.
47. Saglani S, Lui S, Ullmann N, Campbell GA, Sherburn RT, Mathie SA, Denney L, Bossley CJ, Oates T, Walker SA, Bush A, Lloyd CM. IL-33 promotes airway remodeling in pediatric patients with severe steroid-resistant asthma. J Allergy Clin Immunol 2013; 132:676-685.e13.
48. Guo Z, Wu J, Zhao J, Liu F, Chen Y, Bi L, Liu S, Dong L. IL-33 promotes airway remodeling and is a marker of asthma disease severity. J Asthma 2014; 51:863-9.
49. Gregory LG, Jones CP, Walker SA, Sawant D, Gowers KH, Campbell GA, McKenzie AN, Lloyd CM. IL-25 drives remodelling in allergic airways disease induced by house dust mite. Thorax 2013; 68:82-90.
50. Xu X, Luo S, Li B, Dai H, Zhang J. IL-25 contributes to lung fibrosis by directly acting on alveolar epithelial cells and fibroblasts. Exp Biol Med (Maywood). 2019; 244:770-780.
51.Redhu NS, Shan L, Movassagh H, Gounni AS. Thymic stromal lymphopoietin induces migration in human airway smooth muscle cells. Sci Rep 2013; 3:2301.
52.Kaur D, Gomez E, Doe C, Berair R, Woodman L, Saunders R, Hollins F, Rose FR, Amrani Y, May R, Kearley J, Humbles A, Cohen ES, Brightling CE. IL-33 drives airway hyper-responsiveness through IL-13-mediated mast cell: airway smooth muscle crosstalk. Allergy 2015; 70:556-67.
53.Moses HL, Yang EY, Pietenpol JA. Regulation of epithelial proliferation by TGF-beta. Ciba Found Symp 1991; 157:66-74; discussion 75-80.
54.Vignola AM, Chanez P, Chiappara G, Merendino A, Pace E, Rizzo A, la Rocca AM, Bellia V, Bonsignore G, Bousquet J. Transforming growth factor-beta expression in mucosal biopsies in asthma and chronic bronchitis. Am J Respir Crit Care Med. 1997 Aug;156(2 Pt 1):591-9.
55.Brewster CEP, Howarth PH, Djukanovic R, Wilson J, Holgate ST, Roche WR. Myofibroblasts and subepithelial fibrosis in bronchial asthma. Am J Respir Cell Mol Biol 1990; 3:507-11.
56.Wang CH, Huang CD, Lin HC, Lee KY, Lin SM, Liu CY, Huang KH, Ko YS, Chung KF, Kuo HP. Increased circulating fibrocytes in asthma with chronic airflow obstruction. Am J Respir Crit Care Med 2008; 178:583-91.
57.Hoshino M, Nakamura Y, Sim J, Shimojo J, Isogai S. Bronchial subepithelial fibrosis and expression of matrix metalloproteinase-9 in asthmatic airway inflammation. J Allergy Clin Immunol 1998; 102:783-8.
58.Rödel J, Woytas M, Groh A, Schmidt KH, Hartmann M, Lehmann M, Straube E. Production of basic fibroblast growth factor and interleukin 6 by human smooth muscle cells following infection with Chlamydia pneumoniae. Infect Immun 2000; 68:3635-41.
59.John AE, Zhu YM, Brightling CE, Pang L, Knox AJ. Human airway smooth muscle cells from asthmatic individuals have CXCL8 hypersecretion due to increased NF-kappa B p65, C/EBP beta, and RNA polymerase II binding to the CXCL8 promoter. J Immunol 2009; 183:4682-92.
60.Elshaw SR, Henderson N, Knox AJ, Watson SA, Buttle DJ, Johnson SR. Matrix metalloproteinase expression and activity in human airway smooth muscle cells. Br J Pharmacol 2004; 142:1318-24.
61.Benayoun L, Druilhe A, Dombret MC, Aubier M, Pretolani M. Airway structural alterations selectively associated with severe asthma. Am J Respir Crit Care Med 2003; 167:1360-8.
62.Cockcroft DW, Davis BE. Mechanisms of airway hyperresponsiveness. J Allergy Clin Immunol 2006; 118:551-9; quiz 560-1.
63.Bara I, Ozier A, Tunon de Lara JM, Marthan R, Berger P. Pathophysiology of bronchial smooth muscle remodelling in asthma. Eur Respir J 2010; 36:1174-84.
64.Xiong DJP, Martin JG, Lauzon AM. Airway smooth muscle function in asthma. Front Physiol 2022; 13:993406.
65.Salvato G. Quantitative and morphological analysis of the vascular bed in bronchial biopsy specimens from asthmatic and non-asthmatic subjects. Thorax 2001; 56:902-6.
66.Vrugt B, Wilson S, Bron A, Holgate ST , Djukanovic R, Aalbers R. Bronchial angiogenesis in severe glucocorticoid dependent asthma. Eur Respir J 2000; 15:1014-21.
67.Brewster CEP, Howarth PH, Djukanovic R, Wilson J, Holgate ST, Roche WR. Myofibroblasts and subepithelial fibrosis in bronchial asthma. Am J Respir Cell Mol Biol 1990; 3:507-11.
68.Zhang S, Mohammed Q, Burbridge A, Morland CM, Roche WR. Cell cultures from bronchial subepithelial myofibroblasts enhance eosinophil survival in vitro. Eur Respir J 1996; 9:1839-46.
69.Minshall EM, Leung DYM, Martin RJ et al. Eosinophil-associated TGF- ß1 mRNA expression and airways fibrosis in bronchial asthma. Am J Respir Cell Mol Biol 1997; 17:326-33.
70.Xu YD, Hua J, Mui A, O’Connor R, Grotendorst G, Khalil N. Release of biological active TGF-β1 by alveolar epithelial cells results in pulmonary fibrosis. Am J Physiol Lung Cell Mol Physiol 2003; 285: L527–L539.
71.
Perng DW, Wu YC, Chang KT, Wu MT, Chiou YC, Su KC, Perng RP, Lee YC. Leukotriene C4 induces TGF-β1 production in airway epithelium via p38 kinase pathway. Am J Respir Cell Mol Biol 2006; 34:101-7.
72.Kaminska B, Wesolowska A, Danilkiewicz M. TGF-beta signaling and its role in tumor pathogenesis. Acta Biochim Pol 2005; 52: 329–337.
73.Chu HW, Balzar S, Seedorf GJ, Westcott JY, Trudeau JB, Silkoff P, Wenzel SE. Transforming growth factor-β2 induces bronchial epithelial mucin expression in asthma. Am J Pathol 2004; 165:1097-106.
74.McMillan SJ, Xanthon G, Lloyd CM. Manipulation of allergen-induced airway remodeling by treatment with anti-TGF-β antibody effect on the Smad signaling pathway. J Immunol 2005; 174: 5774–5780.
75.Leivonen SK, Häkkinen L, Liu D, Kähäri VM. Smad3 and extracellular signal-regulated kinase 1/2 coordinately mediate transforming growth factor-β-induced expression of connective tissue growth factor in human fibroblasts. J Invest Dermatol 2005; 124:1162-9.
76.Samarakoon R, Higgins CE, Higgins SP, Kutz SM, Higgins PJ. Plasminogen activator inhibitor type-1 gene expression and induced migration in TGF-β 1-stimulated smooth muscle cells is pp60(csrc)/MEK-dependent. J Cell Physiol 2005; 204:236–246.
77.Yue J, Mulder KM. Requirement of Ras/MAPK pathway activation by transforming growth factor b for transforming growth factor β1 production in a Smad-dependent pathway. J Biol Chem 2000; 275: 30765–30773.
78.Wen FQ, Liu X, Kobayashi T, Abe S, Fang Q, Kohyama T, Ertl R, Terasaki Y, Manouilova L, Rennard SI. Interferon-gamma inhibits transforming growth factor-beta production in human airway epithelial cells by targeting Smads. Am J Respir Cell Mol Biol 2004; 30:816-22.
79.Ignotz RA, Endo T, Massague J. Regulation of fribronectin ans type I collagen mRNA levels by transforming growth factor-beta. J Biol Chem 1987; 262:6443-46.
80.Massague J. The transforming growth factor-beta family. Annu Rev Cell Biol 1990; 6:597-641.
81.Kovacs EJ, DiPietro LA. Fibrogenic cytokines and connective tissue production. FASEB J 1994; 8:854-61.
82.Redlich CA, Delisser HM, Elias JA. Retinoic acid inhibition of transforming growth factor-beta-induced collagen production by human lung fibroblasts. Am J Respir Cell Mol Biol 1995; 12:287-95.
83.Morishima Y, Nomura A, Uchida Y et al. Triggering the induction of myofibroblast and fibrogenesis by airway epithelial shedding. Am J Respir Cell Mol Biol 2001; 24:1-11.
84.Makinde T, Murphy RF, Agrawal DK. The regulatory role of TGF-beta in airway remodeling in asthma. Immunol Cell Biol 2007; 85:348-56.
85.Kocwin M, Jonakowski M, Przemecka M, Ziolo J, Panek M, Kuna P. The role of the TGF-SMAD signalling pathway in the etiopathogenesis of severe asthma. Pneumonol Alergol Pol 2016; 84:290-301.
86.Amishima M, Munakata M, Nasuhara Y et al. Expression of epidermal growth factor and epidermal growth factor receptor immunoreactivity in the asthmatic human airway. Am J Respir Crit Care Med 1998; 157:1907-12.
87.Puddicombe SM, Torres-Lozano C, Richter A, Bucchieri F, Lordan JL, Howarth PH, Vrugt B, Albers R, Djukanovic R, Holgate ST, Wilson SJ, Davies DE. Increased expression of p21(waf) cyclin-dependent kinase inhibitor in asthmatic bronchial epithelium. Am J Respir Cell Mol Biol 2003; 28:61-8.
88.Chung KF, Barnes PJ. Cytokines in asthma. Thorax 1999; 54:825-57.
89.Ollerenshaw S, Jarvis D, Sullivan C et al. Substance P immunoreactive nerves in airways and non-asthmatics. Eur Respir J 1991; 4:673-82.
90.Ollerenshaw J, Jarvis D, Woolcock A et al. Absence of immunoreactive vasoactive intestinal polypeptide in tissue from lungs of patients with asthma. N Engl J Med 1989; 320:1244-48.
91.Howarth P, Springall D, Redington A et al. Neuro-peptide-containing nerves in endobronchial biopses from asthmatic and non-asthmatic subjects. Am J Respir Cell Mol Biol 1995; 13:288-96.
92.Poto R, Criscuolo G, Marone G, Brightling CE, Varricchi G. Human Lung Mast Cells: Therapeutic Implications in Asthma. Int J Mol Sci 2022; 23:14466.
93.Ali Komi ED, Bjermer L. Mast cell-mediated orchestration of the immune responses in human allergic asthma: current insights. Clin Rev Allergy Immunol 2019; 56:234–47.
94.Chetta A, Zanini A, Foresi A, et al. Vascular component of airway remodeling in asthma is reduced by high dose of fluticasone. Am J Respir Crit Care Med 2003; 167:751–757.
95.Hoshino M, Takahashi M, Aoike N. Expression of vascular endothelial growth factor, basic fibroblast growth factor, and angiogenin immunoreactivity in asthmatic airways and its relationship to angiogenesis. J Allergy Clin Immunol 2001; 107:295–301.
96.van der Veen TA, de Groot LES, Melgert BN. The different faces of the macrophage in asthma. Curr Opin Pulm Med 2020; 26:62-68.
97.Balhara J, Gounni AS. The alveolar macrophages in asthma: a double-edged sword. Mucosal Immunol 2012; 5: 605–609.
98.Braile M, Fiorelli A, Sorriento D, Di Crescenzo RM, Galdiero MR, Marone G, Santini M, Varricchi G, Loffredo S. Human Lung-Resident Macrophages Express and Are Targets of Thymic Stromal Lymphopoietin in the Tumor Microenvironment. Cells 2021 6; 10:2012.